Enhancing Photosynthesis
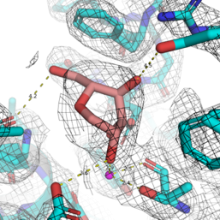
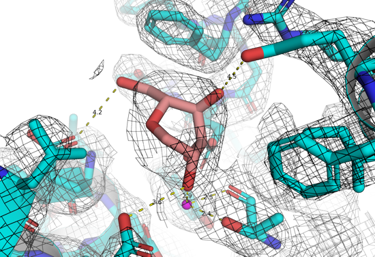
La fotosintesi è il processo mediante il quale la luce solare è convertita in potenziale chimico, utilizzabile dagli organismi, e rappresenta pertantp il sito primario si produzione energetica nella biosfera. In condizioni ottimali le relazioni di foto-conversione procedono con una resa quantica estremamente elevata (80-99%). Lo studio di questi sistemi biologici può quindi rappresentate un paradigma utile per l’implementazione di materiali e strategie che li imitino per applicazioni fotovoltaiche e fotocatalitiche. D’altro canto, in condizioni naturali, a causa di variabilità e fluttuazioni ambientali, la resa fotosintetica è generalmente più bassa di quella ottimale e questo può portare a una riduzione della produttività, soprattutto per quanto riguarda le colture agricole. Pertanto, lo studio dei processi, come la raccolta della luce e il controllo dell’efficienza di raccolta, potrebbe portare ad un miglioramento della produttività delle colture ed essere quindi vantaggioso dal punto di vista sociale.
Il progetto “Enhancing Photosintesi” si propone di affrontare alcuni di questi aspetti, studiando sia i fattori che influenzano la produttività delle colture/piante, sia i loro meccanismi molecolari, sia i meccanismi fondamentali di conversione dei fotoni nei fotosistemi con l’obiettivo di trasferire alcune delle loro caratteristiche in sistemi biomimetici artificiali.
Il progetto prevede inoltre lo sviluppo di una piattaforma spettroscopica (ottica) per lo studio dei processi fotosintetici sia in(super)complessi clorofilla-proteina isolati, sia in sistemi fotosintetici intatti così come in sistemi sintetici artificiali (anche allo stato solido). La strumentazione coprirà oltre ad un ampio intervallo temporale (dai femto ai millisecondi) anche una ambia banda spettrale (dal vicino UV al vicino IR). Questa piattaforma sarà basata sia su strumentazioni già esistenti presso il CNR Milano e al PoliMI, le cui caratteristiche saranno sostanzialmente estese/implementate per renderle adatte allo studio di campioni molto diversificati e complicati da studiare a causa dell’elevata dispersione della luce che presentano. E’ fine del progetto rendere questo “hub” spettroscopico aperto che rappresenti una struttura di duratura che possa aggregare e promuovere la ricerca in fotosintesi, e su argomenti strettamente correlati, a livello regionale e possibilmente nazionale.